






The CRG is an international biomedical research institute of excellence...
More information
Sistema CERCA

Excelencia Severo Ochoa

HR Excellence in research
![]()
European Union





 News
News
15
Jul

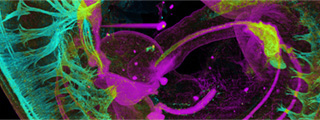
A study in Nature produced the most complete atlas to date of the DNA regions that switch human genes on and off
14
Jul

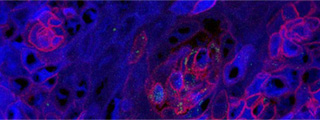
The CRG is one of ten centres recognised in this round.
13
Jul

The CRG's new director on the questions driving her science, the kind of institute she wants to build and widening who gets to do research
 Events
Events
14
Sep

“ELIXIR course: Artificial Intelligence and Machine Learning in Life Sciences” focuses on acquiring the conceptual understanding and practical skills required to apply ML methods appropriately, critically, and reproducibly. The course targets...
22
Sep

La Alianza de Centros de Excelencia Severo Ochoa y Unidades de Excelencia María de Maeztu (SOMMa) junto al Centro de Regulación Genómica (CRG) organizan el evento SOMMa Connect que tendrá lugar el 22 y 23 de septiembre de 2026 en el auditorio del...
28
Sep

The symposium follows the collaborative spirit of the Barcelona Collaboratorium, combining scientific excellence with strong integration of early-career researchers through discussion sessions, networking opportunities, and community-building...
BOARD OF TRUSTEES
Generalitat de Catalunya
![]()
Ministerio de Ciencia e Innovación

Universitat Pompeu Fabra
![]()
"la Caixa Foundation"
MEMBERS OF
The Barcelona Institute of Science and Technology
![]()
SOMMa

Barcelona Biomedical Research Park
![]()
EU Life

Core for life

AseBio
